Can You Complete These Sentences That Describe How a Cell Performs Work?
Affiliate 4: Introduction to How Cells Obtain Energy
iv.1 Energy and Metabolism
Learning Objectives
Past the end of this section, you will be able to:
- Explain what metabolic pathways are
- State the start and second laws of thermodynamics
- Explain the difference betwixt kinetic and potential energy
- Describe endergonic and exergonic reactions
- Discuss how enzymes function as molecular catalysts
Watch a video about heterotrophs.
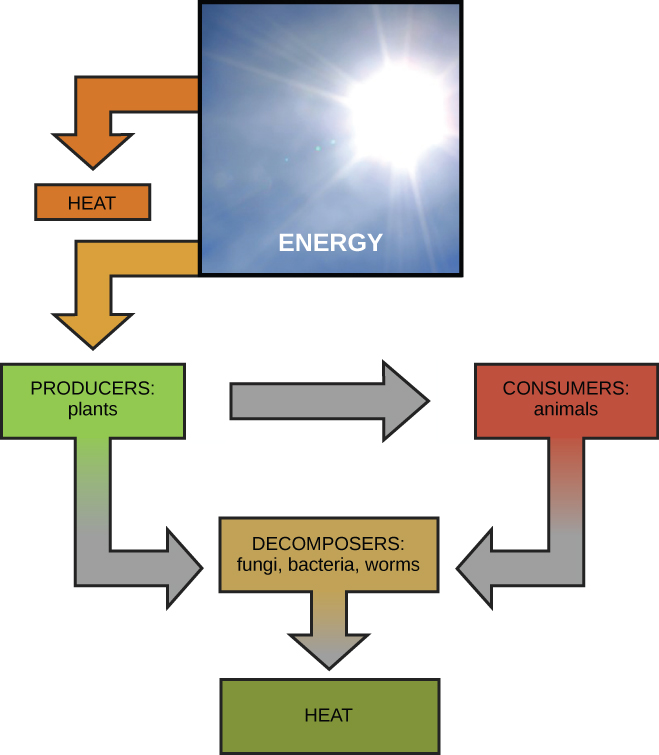
Scientists use the term bioenergetics to draw the concept of energy flow (Figure 4.2) through living systems, such as cells. Cellular processes such as the edifice and breaking downwardly of complex molecules occur through stepwise chemic reactions. Some of these chemic reactions are spontaneous and release energy, whereas others crave energy to keep. Just equally living things must continually eat food to replenish their energy supplies, cells must continually produce more energy to replenish that used past the many energy-requiring chemical reactions that constantly take place. Together, all of the chemical reactions that take place inside cells, including those that consume or generate energy, are referred to as the cell's metabolism.
Metabolic Pathways
Consider the metabolism of carbohydrate. This is a classic case of i of the many cellular processes that apply and produce energy. Living things consume sugars as a major free energy source, because sugar molecules have a great deal of energy stored inside their bonds. For the almost function, photosynthesizing organisms like plants produce these sugars. During photosynthesis, plants use energy (originally from sunlight) to convert carbon dioxide gas (CO2) into sugar molecules (like glucose: C6H12O6). They eat carbon dioxide and produce oxygen as a waste product. This reaction is summarized equally:
6COii + 6H2O + energy ——-> Chalf-dozenH12Ohalf dozen+ 6O2
Because this process involves synthesizing an energy-storing molecule, it requires energy input to proceed. During the light reactions of photosynthesis, free energy is provided by a molecule called adenosine triphosphate (ATP), which is the chief free energy currency of all cells. Just every bit the dollar is used as currency to buy goods, cells use molecules of ATP as energy currency to perform immediate work. In dissimilarity, energy-storage molecules such every bit glucose are consumed only to exist broken down to use their free energy. The reaction that harvests the energy of a sugar molecule in cells requiring oxygen to survive can exist summarized by the opposite reaction to photosynthesis. In this reaction, oxygen is consumed and carbon dioxide is released as a waste product. The reaction is summarized as:
Chalf dozenH12Osix + 6O2 ——> 6CO2 + 6H2O + energy
Both of these reactions involve many steps.
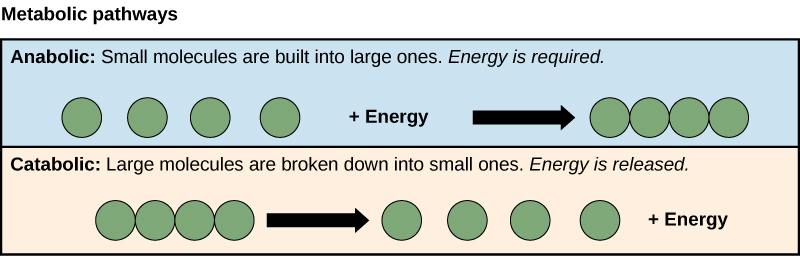
The processes of making and breaking downwardly saccharide molecules illustrate two examples of metabolic pathways. A metabolic pathway is a serial of chemical reactions that takes a starting molecule and modifies it, step-by-step, through a series of metabolic intermediates, eventually yielding a terminal product. In the example of sugar metabolism, the kickoff metabolic pathway synthesized saccharide from smaller molecules, and the other pathway broke saccharide down into smaller molecules. These two opposite processes—the showtime requiring energy and the second producing energy—are referred to as anabolic pathways (edifice polymers) and catabolic pathways (breaking downwards polymers into their monomers), respectively. Consequently, metabolism is composed of synthesis (anabolism) and deposition (catabolism) (Figure 4.iii).
It is of import to know that the chemical reactions of metabolic pathways practice not accept identify on their own. Each reaction step is facilitated, or catalyzed, by a poly peptide called an enzyme. Enzymes are important for catalyzing all types of biological reactions—those that crave free energy as well as those that release free energy.
Energy
Thermodynamics refers to the study of energy and energy transfer involving physical matter. The matter relevant to a detail example of free energy transfer is chosen a system, and everything exterior of that matter is chosen the environment. For instance, when heating a pot of water on the stove, the system includes the stove, the pot, and the water. Free energy is transferred inside the organization (between the stove, pot, and h2o). There are 2 types of systems: open and airtight. In an open arrangement, energy tin be exchanged with its environment. The stovetop organization is open considering rut can be lost to the air. A airtight organization cannot exchange energy with its environs.
Biological organisms are open up systems. Energy is exchanged between them and their surroundings as they apply free energy from the lord's day to perform photosynthesis or eat energy-storing molecules and release energy to the environment by doing work and releasing oestrus. Like all things in the physical world, energy is bailiwick to physical laws. The laws of thermodynamics govern the transfer of energy in and among all systems in the universe.
In general, free energy is defined as the power to do work, or to create some kind of alter. Energy exists in different forms. For instance, electrical energy, light energy, and heat free energy are all unlike types of energy. To appreciate the style free energy flows into and out of biological systems, it is important to sympathise two of the physical laws that govern free energy.
Thermodynamics
The kickoff police force of thermodynamics states that the total amount of energy in the universe is abiding and conserved. In other words, at that place has always been, and e'er will be, exactly the same amount of energy in the universe. Energy exists in many different forms. According to the get-go constabulary of thermodynamics, energy may be transferred from place to place or transformed into different forms, but it cannot be created or destroyed. The transfers and transformations of energy have identify around us all the time. Calorie-free bulbs transform electrical energy into lite and heat energy. Gas stoves transform chemical energy from natural gas into estrus free energy. Plants perform ane of the most biologically useful energy transformations on world: that of converting the energy of sunlight to chemic energy stored within organic molecules (Figure four.2). Some examples of energy transformations are shown in Figure iv.4.
The challenge for all living organisms is to obtain energy from their surround in forms that they can transfer or transform into usable energy to practise work. Living cells have evolved to meet this challenge. Chemical energy stored within organic molecules such as sugars and fats is transferred and transformed through a serial of cellular chemic reactions into free energy within molecules of ATP. Energy in ATP molecules is easily accessible to do work. Examples of the types of piece of work that cells demand to do include building complex molecules, transporting materials, powering the motion of cilia or flagella, and contracting musculus fibers to create motility.

A living prison cell'south primary tasks of obtaining, transforming, and using free energy to exercise work may seem unproblematic. Still, the 2nd law of thermodynamics explains why these tasks are harder than they appear. All energy transfers and transformations are never completely efficient. In every energy transfer, some corporeality of energy is lost in a form that is unusable. In most cases, this course is heat free energy. Thermodynamically, oestrus free energy is defined every bit the energy transferred from ane organisation to some other that is non work. For example, when a calorie-free seedling is turned on, some of the energy being converted from electric energy into light free energy is lost every bit oestrus energy. As well, some energy is lost as rut energy during cellular metabolic reactions.
An important concept in physical systems is that of order and disorder. The more than free energy that is lost by a system to its surroundings, the less ordered and more than random the arrangement is. Scientists refer to the measure of randomness or disorder within a organization as entropy. High entropy ways high disorder and low energy. Molecules and chemic reactions have varying entropy also. For example, entropy increases as molecules at a high concentration in one identify diffuse and spread out. The 2d police force of thermodynamics says that energy volition always be lost equally rut in free energy transfers or transformations.
Living things are highly ordered, requiring constant energy input to be maintained in a state of low entropy.
Potential and Kinetic Free energy
When an object is in motion, there is energy associated with that object. Think of a wrecking ball. Even a slow-moving wrecking brawl can do a great deal of damage to other objects. Free energy associated with objects in motion is called kinetic energy (Figure 4.5). A speeding bullet, a walking person, and the rapid movement of molecules in the air (which produces heat) all have kinetic energy.
Now what if that same motionless wrecking ball is lifted two stories in a higher place ground with a crane? If the suspended wrecking ball is unmoving, is in that location energy associated with it? The answer is yes. The energy that was required to lift the wrecking ball did not disappear, only is now stored in the wrecking ball past virtue of its position and the force of gravity acting on it. This type of energy is chosen potential energy (Figure 4.5). If the ball were to fall, the potential energy would be transformed into kinetic energy until all of the potential energy was exhausted when the ball rested on the ground. Wrecking balls likewise swing like a pendulum; through the swing, at that place is a constant modify of potential energy (highest at the top of the swing) to kinetic energy (highest at the lesser of the swing). Other examples of potential energy include the energy of h2o held behind a dam or a person most to skydive out of an airplane.

Potential energy is not merely associated with the location of matter, simply also with the structure of matter. Fifty-fifty a spring on the ground has potential energy if it is compressed; then does a rubber band that is pulled taut. On a molecular level, the bonds that hold the atoms of molecules together exist in a particular structure that has potential energy. Remember that anabolic cellular pathways require energy to synthesize complex molecules from simpler ones and catabolic pathways release free energy when complex molecules are broken downwards. The fact that energy can be released by the breakup of certain chemical bonds implies that those bonds have potential energy. In fact, there is potential energy stored inside the bonds of all the food molecules we swallow, which is eventually harnessed for use. This is because these bonds can release energy when cleaved. The type of potential free energy that exists within chemic bonds, and is released when those bonds are cleaved, is called chemical free energy. Chemical free energy is responsible for providing living cells with energy from food. The release of energy occurs when the molecular bonds within nutrient molecules are cleaved.
Sentry a video virtually kilocalories.
Concept in Activeness

Visit the site and select "Pendulum" from the "Work and Energy" bill of fare to run into the shifting kinetic and potential energy of a pendulum in motion.
Free and Activation Free energy
After learning that chemical reactions release energy when energy-storing bonds are broken, an important side by side question is the following: How is the energy associated with these chemical reactions quantified and expressed? How can the free energy released from 1 reaction exist compared to that of another reaction? A measurement of costless free energy is used to quantify these energy transfers. Retrieve that co-ordinate to the 2d constabulary of thermodynamics, all free energy transfers involve the loss of some amount of energy in an unusable form such as rut. Free energy specifically refers to the energy associated with a chemical reaction that is available subsequently the losses are accounted for. In other words, energy is usable energy, or energy that is available to practice work.
If energy is released during a chemical reaction, and so the change in complimentary energy, signified as ∆G (delta G) will be a negative number. A negative change in free energy also means that the products of the reaction take less gratuitous free energy than the reactants, because they release some free energy during the reaction. Reactions that have a negative modify in free energy and consequently release energy are called exergonic reactions. Think: exergonic ways energy is exiting the system. These reactions are also referred to every bit spontaneous reactions, and their products take less stored energy than the reactants. An of import distinction must be drawn between the term spontaneous and the idea of a chemical reaction occurring immediately. Opposite to the everyday use of the term, a spontaneous reaction is not one that all of a sudden or quickly occurs. The rusting of iron is an example of a spontaneous reaction that occurs slowly, little by piffling, over time.
If a chemical reaction absorbs energy rather than releases energy on balance, then the ∆G for that reaction will exist a positive value. In this case, the products have more free energy than the reactants. Thus, the products of these reactions can be idea of equally energy-storing molecules. These chemical reactions are called endergonic reactions and they are non-spontaneous. An endergonic reaction will not take place on its own without the addition of costless free energy.

Wait at each of the processes shown and make up one's mind if it is endergonic or exergonic.
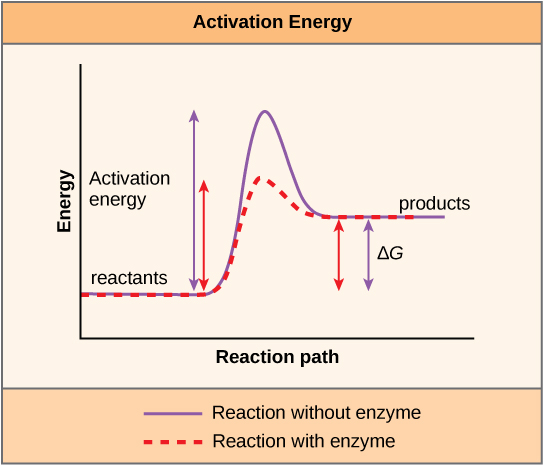
At that place is another important concept that must be considered regarding endergonic and exergonic reactions. Exergonic reactions crave a small amount of energy input to get going, before they tin keep with their free energy-releasing steps. These reactions accept a net release of energy, but still require some energy input in the offset. This small corporeality of energy input necessary for all chemical reactions to occur is called the activation energy.
Concept in Action

Watch an animation of the move from free free energy to transition land of the reaction.
Enzymes
A substance that helps a chemic reaction to occur is called a catalyst, and the molecules that catalyze biochemical reactions are called enzymes. Most enzymes are proteins and perform the critical task of lowering the activation energies of chemical reactions inside the jail cell. Most of the reactions disquisitional to a living cell happen too slowly at normal temperatures to exist of whatever use to the prison cell. Without enzymes to speed up these reactions, life could not persist. Enzymes exercise this by binding to the reactant molecules and holding them in such a fashion as to make the chemic bond-breaking and -forming processes take place more than easily. It is important to remember that enzymes exercise non change whether a reaction is exergonic (spontaneous) or endergonic. This is considering they do not change the gratuitous energy of the reactants or products. They but reduce the activation energy required for the reaction to go forrard (Figure 4.7). In add-on, an enzyme itself is unchanged past the reaction it catalyzes. One time one reaction has been catalyzed, the enzyme is able to participate in other reactions.
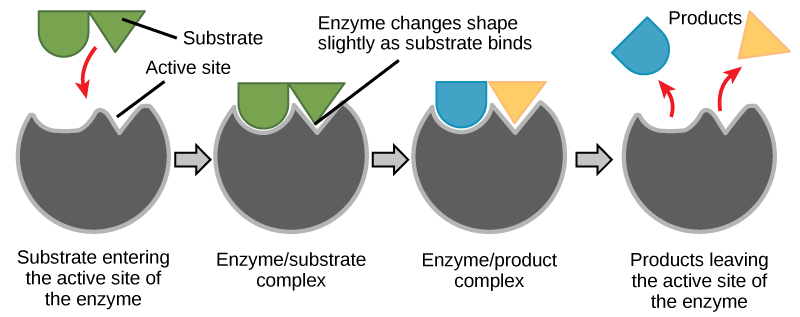
The chemical reactants to which an enzyme binds are called the enzyme'due south substrates. There may be i or more substrates, depending on the item chemical reaction. In some reactions, a single reactant substrate is cleaved down into multiple products. In others, two substrates may come together to create one larger molecule. Two reactants might likewise enter a reaction and both become modified, but they leave the reaction as two products. The location within the enzyme where the substrate binds is chosen the enzyme's active site. The active site is where the "action" happens. Since enzymes are proteins, at that place is a unique combination of amino acid side chains inside the active site. Each side chain is characterized past dissimilar backdrop. They can be big or small, weakly acidic or basic, hydrophilic or hydrophobic, positively or negatively charged, or neutral. The unique combination of side chains creates a very specific chemical environment inside the active site. This specific surroundings is suited to bind to i specific chemical substrate (or substrates).
Active sites are subject to influences of the local environment. Increasing the environmental temperature mostly increases reaction rates, enzyme-catalyzed or otherwise. All the same, temperatures outside of an optimal range reduce the rate at which an enzyme catalyzes a reaction. Hot temperatures volition eventually cause enzymes to denature, an irreversible change in the three-dimensional shape and therefore the office of the enzyme. Enzymes are likewise suited to function all-time inside a certain pH and salt concentration range, and, equally with temperature, extreme pH, and common salt concentrations tin cause enzymes to denature.
For many years, scientists thought that enzyme-substrate binding took place in a simple "lock and key" manner. This model asserted that the enzyme and substrate fit together perfectly in one instantaneous stride. Nevertheless, current research supports a model called induced fit (Effigy 4.8). The induced-fit model expands on the lock-and-key model past describing a more than dynamic binding between enzyme and substrate. As the enzyme and substrate come together, their interaction causes a mild shift in the enzyme's structure that forms an ideal bounden arrangement betwixt enzyme and substrate.
Concept in Action

View an animation of induced fit.
When an enzyme binds its substrate, an enzyme-substrate complex is formed. This complex lowers the activation energy of the reaction and promotes its rapid progression in one of multiple possible ways. On a bones level, enzymes promote chemical reactions that involve more than than one substrate past bringing the substrates together in an optimal orientation for reaction. Another way in which enzymes promote the reaction of their substrates is by creating an optimal environment within the active site for the reaction to occur. The chemic backdrop that emerge from the particular system of amino acid R groups inside an agile site create the perfect environs for an enzyme'south specific substrates to react.
The enzyme-substrate circuitous tin can also lower activation energy by compromising the bond structure so that it is easier to break. Finally, enzymes tin can as well lower activation energies past taking part in the chemical reaction itself. In these cases, it is important to remember that the enzyme will always return to its original country by the completion of the reaction. I of the authentication properties of enzymes is that they remain ultimately unchanged by the reactions they catalyze. After an enzyme has catalyzed a reaction, it releases its product(s) and can catalyze a new reaction.
It would seem ideal to accept a scenario in which all of an organism's enzymes existed in abundant supply and functioned optimally nether all cellular conditions, in all cells, at all times. Nevertheless, a diverseness of mechanisms ensures that this does not happen. Cellular needs and atmospheric condition constantly vary from prison cell to cell, and change within individual cells over time. The required enzymes of stomach cells differ from those of fat storage cells, skin cells, blood cells, and nerve cells. Furthermore, a digestive organ cell works much harder to process and suspension down nutrients during the time that closely follows a repast compared with many hours after a repast. As these cellular demands and atmospheric condition vary, so must the amounts and functionality of unlike enzymes.
Since the rates of biochemical reactions are controlled past activation energy, and enzymes lower and make up one's mind activation energies for chemical reactions, the relative amounts and operation of the variety of enzymes within a cell ultimately determine which reactions will go along and at what rates. This determination is tightly controlled in cells. In certain cellular environments, enzyme activity is partly controlled by environmental factors like pH, temperature, salt concentration, and, in some cases, cofactors or coenzymes.
Enzymes can besides be regulated in ways that either promote or reduce enzyme activity. There are many kinds of molecules that inhibit or promote enzyme function, and various mechanisms by which they do so. In some cases of enzyme inhibition, an inhibitor molecule is similar plenty to a substrate that it can bind to the agile site and simply block the substrate from binding. When this happens, the enzyme is inhibited through competitive inhibition, because an inhibitor molecule competes with the substrate for binding to the active site.
On the other hand, in noncompetitive inhibition, an inhibitor molecule binds to the enzyme in a location other than the active site, chosen an allosteric site, only nonetheless manages to block substrate binding to the active site. Some inhibitor molecules bind to enzymes in a location where their binding induces a conformational change that reduces the affinity of the enzyme for its substrate. This type of inhibition is chosen allosteric inhibition (Figure 4.9). Almost allosterically regulated enzymes are made up of more than one polypeptide, meaning that they have more than one poly peptide subunit. When an allosteric inhibitor binds to a region on an enzyme, all active sites on the poly peptide subunits are changed slightly such that they bind their substrates with less efficiency. There are allosteric activators likewise as inhibitors. Allosteric activators demark to locations on an enzyme away from the agile site, inducing a conformational change that increases the affinity of the enzyme's active site(s) for its substrate(s) (Figure 4.9).
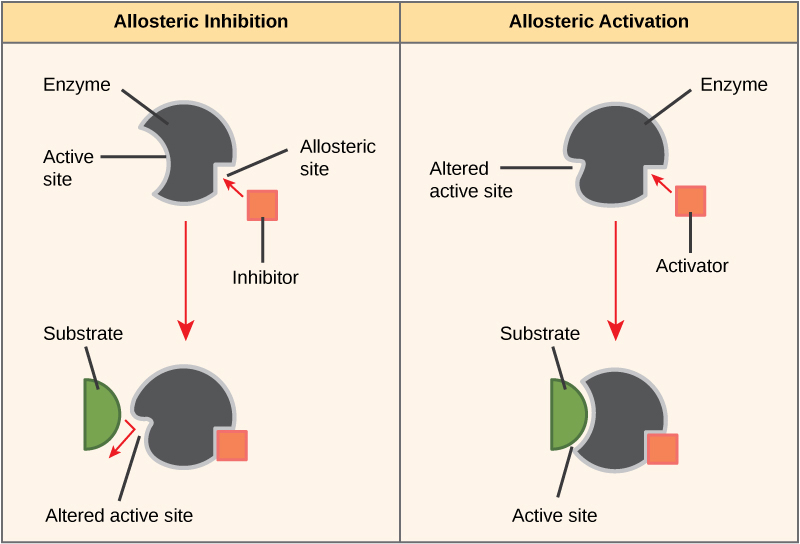
Through the Ethnic Lens
Plants cannot run or hide from their predators and have evolved many strategies to deter those who would eat them. Think of thorns, irritants and secondary metabolites: these are compounds that do not directly help the plant grow, but are made specifically to keep predators away. Secondary metabolites are the most common mode plants deter predators. Some examples of secondary metabolites are atropine, nicotine, THC and caffeine. Humans accept plant these secondary metabolite compounds a rich source of materials for medicines. It is estimated that 90% of the drugs in the modern chemist's have their "roots" in these secondary metabolites.
First peoples herbal treatments revealed these secondary metabolites to the earth. For case, Indigenous peoples take long used the bawl of willow shrubs and alder trees for a tea, tonic or poultice to reduce inflammation. You lot will learn more most the inflammation response past the immune system in affiliate 11.

Both willow and alder bark contain the compound salicin. Nigh of the states have this compound in our medicine closet in the form of salicylic acid or aspirin. Aspirin has been proved to reduce pain and inflammation, and one time in our cells salicin converts to salicylic acrid.
And so how does it work? Salicin or aspirin acts as an enzyme inhibitor. In the inflammatory response two enzymes, COX1 and COX2 are key to this process. Salicin or aspirin specifically modifies an amino acid (serine) in the agile site of these two related enzymes. This modification of the active sites does non permit the normal substrate to demark and and then the inflammatory process is disrupted. As yous have read in this affiliate, this makes it competitive enzyme inhibitor.
Pharmaceutical Drug Developer

Enzymes are key components of metabolic pathways. Agreement how enzymes work and how they can exist regulated are key principles behind the evolution of many of the pharmaceutical drugs on the market place today. Biologists working in this field interact with other scientists to pattern drugs (Figure 4.eleven).
Consider statins for example—statins is the proper noun given to i class of drugs that can reduce cholesterol levels. These compounds are inhibitors of the enzyme HMG-CoA reductase, which is the enzyme that synthesizes cholesterol from lipids in the trunk. By inhibiting this enzyme, the level of cholesterol synthesized in the body tin can exist reduced. Similarly, acetaminophen, popularly marketed under the brand name Tylenol, is an inhibitor of the enzyme cyclooxygenase. While information technology is used to provide relief from fever and inflammation (hurting), its mechanism of activeness is yet not completely understood.
How are drugs discovered? One of the biggest challenges in drug discovery is identifying a drug target. A drug target is a molecule that is literally the target of the drug. In the case of statins, HMG-CoA reductase is the drug target. Drug targets are identified through painstaking enquiry in the laboratory. Identifying the target lonely is non enough; scientists also need to know how the target acts inside the cell and which reactions become amiss in the case of illness. Once the target and the pathway are identified, then the actual process of drug design begins. In this stage, chemists and biologists work together to design and synthesize molecules that can block or activate a detail reaction. Still, this is but the beginning: If and when a drug image is successful in performing its function, and then it is subjected to many tests from in vitro experiments to clinical trials before it tin can get approval from the U.S. Nutrient and Drug Administration to be on the market.
Many enzymes exercise non work optimally, or fifty-fifty at all, unless bound to other specific not-poly peptide helper molecules. They may bail either temporarily through ionic or hydrogen bonds, or permanently through stronger covalent bonds. Bounden to these molecules promotes optimal shape and function of their respective enzymes. Two examples of these types of helper molecules are cofactors and coenzymes. Cofactors are inorganic ions such every bit ions of iron and magnesium. Coenzymes are organic helper molecules, those with a basic diminutive structure made up of carbon and hydrogen. Like enzymes, these molecules participate in reactions without being changed themselves and are ultimately recycled and reused. Vitamins are the source of coenzymes. Some vitamins are the precursors of coenzymes and others act directly as coenzymes. Vitamin C is a direct coenzyme for multiple enzymes that take part in building the important connective tissue, collagen. Therefore, enzyme role is, in function, regulated by the abundance of various cofactors and coenzymes, which may be supplied by an organism's diet or, in some cases, produced by the organism.
Feedback Inhibition in Metabolic Pathways
Molecules can regulate enzyme office in many means. The major question remains, however: What are these molecules and where do they come from? Some are cofactors and coenzymes, every bit you lot have learned. What other molecules in the cell provide enzymatic regulation such every bit allosteric modulation, and competitive and non-competitive inhibition? Perhaps the virtually relevant sources of regulatory molecules, with respect to enzymatic cellular metabolism, are the products of the cellular metabolic reactions themselves. In a most efficient and elegant manner, cells have evolved to use the products of their own reactions for feedback inhibition of enzyme action. Feedback inhibition involves the employ of a reaction production to regulate its ain further production (Figure 4.12). The cell responds to an abundance of the products by slowing down production during anabolic or catabolic reactions. Such reaction products may inhibit the enzymes that catalyzed their production through the mechanisms described above.
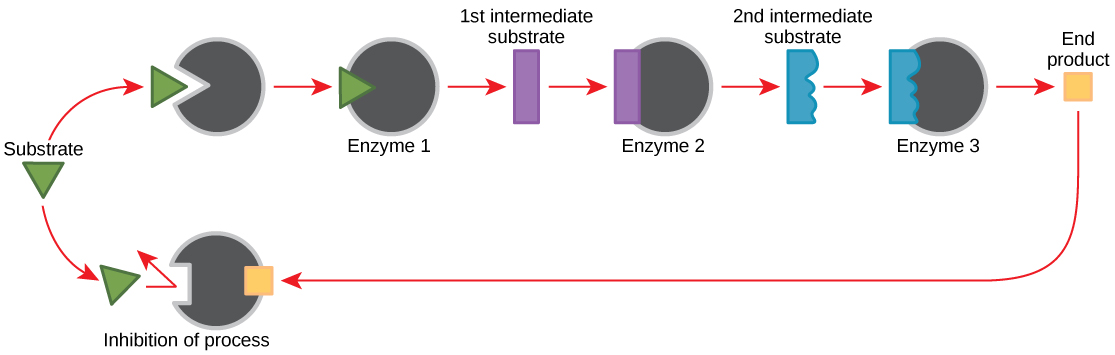
The production of both amino acids and nucleotides is controlled through feedback inhibition. Additionally, ATP is an allosteric regulator of some of the enzymes involved in the catabolic breakdown of sugar, the process that creates ATP. In this mode, when ATP is in arable supply, the cell can prevent the production of ATP. On the other hand, ADP serves equally a positive allosteric regulator (an allosteric activator) for some of the aforementioned enzymes that are inhibited by ATP. Thus, when relative levels of ADP are high compared to ATP, the cell is triggered to produce more ATP through carbohydrate catabolism.
Section Summary
Cells perform the functions of life through various chemical reactions. A prison cell's metabolism refers to the combination of chemic reactions that take place within it. Catabolic reactions break down complex chemicals into simpler ones and are associated with energy release. Anabolic processes build complex molecules out of simpler ones and require energy.
In studying energy, the term system refers to the thing and environment involved in energy transfers. Entropy is a measure of the disorder of a system. The physical laws that describe the transfer of free energy are the laws of thermodynamics. The beginning police force states that the total amount of energy in the universe is constant. The second law of thermodynamics states that every energy transfer involves some loss of energy in an unusable form, such as heat free energy. Energy comes in dissimilar forms: kinetic, potential, and free. The modify in complimentary energy of a reaction can be negative (releases energy, exergonic) or positive (consumes energy, endergonic). All reactions require an initial input of energy to go along, called the activation free energy.
Enzymes are chemical catalysts that speed upwards chemical reactions by lowering their activation energy. Enzymes have an active site with a unique chemical surroundings that fits item chemical reactants for that enzyme, chosen substrates. Enzymes and substrates are idea to bind co-ordinate to an induced-fit model. Enzyme action is regulated to conserve resources and reply optimally to the surroundings.
Glossary
activation energy: the corporeality of initial free energy necessary for reactions to occur
agile site: a specific region on the enzyme where the substrate binds
allosteric inhibition: the mechanism for inhibiting enzyme action in which a regulatory molecule binds to a 2d site (not the active site) and initiates a conformation change in the agile site, preventing binding with the substrate
anabolic: describes the pathway that requires a net free energy input to synthesize circuitous molecules from simpler ones
bioenergetics: the concept of free energy flow through living systems
catabolic: describes the pathway in which complex molecules are broken down into simpler ones, yielding energy as an additional product of the reaction
competitive inhibition: a general mechanism of enzyme activeness regulation in which a molecule other than the enzyme's substrate is able to bind the active site and foreclose the substrate itself from binding, thus inhibiting the overall rate of reaction for the enzyme
endergonic: describes a chemical reaction that results in products that store more than chemical potential energy than the reactants
enzyme: a molecule that catalyzes a biochemical reaction
exergonic: describes a chemical reaction that results in products with less chemical potential energy than the reactants, plus the release of free free energy
feedback inhibition: a mechanism of enzyme activity regulation in which the product of a reaction or the final product of a series of sequential reactions inhibits an enzyme for an before step in the reaction serial
heat energy: the energy transferred from 1 system to some other that is non work
kinetic energy: the type of energy associated with objects in movement
metabolism: all the chemic reactions that take identify inside cells, including those that use energy and those that release energy
noncompetitive inhibition: a general mechanism of enzyme action regulation in which a regulatory molecule binds to a site other than the agile site and prevents the active site from binding the substrate; thus, the inhibitor molecule does not compete with the substrate for the agile site; allosteric inhibition is a course of noncompetitive inhibition
potential energy: the type of energy that refers to the potential to do work
substrate: a molecule on which the enzyme acts
thermodynamics: the scientific discipline of the relationships between oestrus, energy, and work
Source: https://opentextbc.ca/biology/chapter/4-1-energy-and-metabolism/
0 Response to "Can You Complete These Sentences That Describe How a Cell Performs Work?"
Enregistrer un commentaire